Next Section |
Table of Contents
SCIENTIFIC OBJECTIVES
External versus Internal Controls
The apparent coincidence between the occurrence of giant mound clusters and potential deeper-lying hydrocarbon deposits suggests a possible internal control from (mostly transient) fluxes of geofluids from deep geological reservoirs to the seabed. Sedimentary buildup might have been controlled by microbial communities, which also may have played an active role in the stabilization of the steep flanks and in the possible lithification of the (hitherto not yet sampled) mound core through automicrite formation.
On the other hand, these mounds are located on a margin that throughout the Neogene to Quaternary has repeatedly alternated between glacial and interglacial environments. There is also increasing evidence from results of the ACES, ECOMOUND, and GEOMOUND projects that the active mound provinces also occur in oceanographically distinct settings (De Mol et al., 2002; Foubert et al., in press; Van Rooij, 2004; Wheeler et al., in press). These mounds cluster in the highest salinity water and also bathymetrically coincide with the spread of the oxygen minimum zone along the deep continental margin (De Mol et al., 2002; Freiwald et al., 2004). In Porcupine Basin, these specific environmental conditions are provided by the northward flow of Mediterranean Outflow Water at intermediated depths (~700–900 m). Locally enhanced currents associated with mixing and interaction of watermasses featuring a density contrast may be at the origin of contrasting effects such as enhanced fluxes of potential nutrients and low sedimentation rates. Such observations consequently also argue for a complex but equally important external control.
In a way, a central hypothesis to be tested is to what extent mound provinces originate at the crossroads of fluxes of internal (trigger phase) and external (relay phase) origin (Henriet et al., 2002).
Of Mounds and Drifts
The thick drift sediment sheet embedding the mounds holds a high-resolution record of past fluctuations of watermasses and paleocurrents on this section of the North-Atlantic margin. Seismic records of exceptionally high resolution may allow the correlation of this record—once verified by drilling—to the mound growth phases. Correlation of the Porcupine drift record with ODP sites along the Atlantic margin opens perspectives of cross-basin comparisons. Corals within the drill cores will provide information on the paleoceanographic conditions, as already substantiated by the pre-IODP coring results (Marion-Dufresne preparatory coring). Variations in terrigeneous content and organic matter in drift sediments should allow us to trace terrestrial sources and shelf-to-slope sediment pathways. The association of mounds and drifts on this upper continental slope thus will also provide a unique paleoenvironmental record of the Atlantic margin.
Hypotheses to be Tested
The objectives of Expedition 307 can be summarized into four major hypotheses:
• Gas seeps act as a prime trigger for mound genesis—a case for geosphere–biosphere coupling.
• Mound "events" frame into a paleoenvironmental plot—prominent erosional surfaces reflect global oceanographic events.
• Mounds are high-resolution paleoenvironmental recorders.
• The Porcupine mounds are present-day analogs for Phanerozoic reef mounds and mud mounds.
1. Gas seeps act as a prime trigger for mound genesis—a case for geosphere–biosphere coupling.
Only drilling to the base of the mounds will allow verification of to what extent fluids may or may not have played a role in mound genesis and/or growth. The question of source rocks in the region and possible migration routes into the study area has been tackled by the teams of FZ Jülich and GFZ Potsdam, Germany. Two-dimensional basin modeling was used to evaluate the possible link between hydrocarbon leakage and mound growth (PetroMod version 8.0, IES GmbH Jülich). Seismic lines of industrial origin and six exploration wells were used for calibration of the burial and thermal history using vitrinite reflectance, bottom hole temperatures, and apatite fission track data. Modeling results indicate that Jurassic and older source rocks are mature to overmature throughout the basin. Cretaceous strata are immature to mature in the central part of the basin and immature on the flanks. The Tertiary sequence remains immature over the entire basin. Hydrocarbon generation started in Late Cretaceous times for the deepest sequences. Phase separation was modeled to occur during migration at depth ranges between 2000 and 4000 m. Upon phase separation, the migration of the free gas phase dominated over that of oil, such that gas is the main migrating fluid in the shallower intervals. Migration is mainly buoyancy driven and vertical. Only Aptian and Tertiary deltaic layers direct hydrocarbon flow out of the basin. The model predicts a potential focusing of gas migration toward the Belgica mounds area, where a pinchout of Cretaceous and Tertiary layers beneath the mound area is observed. The reconstruction shows that seeping gas may have been available for methanotrophic bacteria and related formation of hardgrounds since the Miocene.
During recent cruises that have studied mud volcanoes and mounds in the Gulf of Cadiz, gases, hydrates, and thick authigenic carbonate crusts were sampled associated with corals, clams, and pogonophores. In mid-2004, the Marion-Dufresne collected cores from juvenile mound sites off Morocco: full of corals from top to bottom, they were characterized by a strong H2S signal and sulfidic mineralizations (Foubert et al., 2005). Analysis of very high resolution seismic data below the Belgica mounds (proposed Site PORC-03A) highlighted within the basal sigmoidal sequences acoustic anomalies (amplitude, instantaneous frequency, and polarity) (Figs. F17, F18, F19) possibly related to low quantities of gas. In light of ODP Leg 182 findings off Australia (Feary, Hine, Malone, et al., 1999), such mound/gas associations deserve attention. Gases and possible low molecular weight hydrocarbons in sediments and pore waters need to be analyzed. The carbon isotope signature (12C/13C) of the organic compounds, both in possible authigenic carbonates and in biological skeletal carbonate, may provide clues to hydrocarbon-derived components. Lithogenesis and possible authigenic carbonate formation will be studied through analysis of major, minor, and trace elements by X-ray diffraction (XRF) and laser ablation–inductively coupled plasma–mass spectroscopy (LA-ICP-MS) to provide a record of responses to external and internal controls of mound formation and growth. Subsamples for microbiological analysis will be aseptically obtained from whole-round core sections acquired from selected depths to test the hypothesis that methane-driven microbial activity catalyzes the formation and/or development of carbonate mounds.
Subsamples will be prepared to quantify total bacterial populations (direct microscopy) and their rates of activity (using 35S- and 14C-labeled compounds) and identify active bacterial populations. Extant bacterial populations will be characterized using classical microscopic methods (albeit adapted for deep biosphere sediments) as well as molecular genetic techniques (i.e., 16S ribosomal ribonucleic acid [rRNA]). Comparison with other deep-biosphere sites should help to evaluate whether bacterial populations that could potentially contribute to carbonate mound formation exist. This will be supplemented by shipboard initiation of enrichments and cultivations of important bacterial populations (most probable number [MPN] method). Bacterial phospholipid fatty acid (PLFA) biomarkers should provide complementary evidence of present and past bacterial populations. Furthermore, direct measurements of bacterial methanogenesis and sulfate reduction activity can be used to document ongoing bacterial activity that may be critical to mound formation through the formation of methane via methanogenesis and bicarbonate alkalinity through sulfate reduction. Samples will be taken for pore water analysis and solid-phase organic carbon analysis from the same whole-round cores as for microbiological analysis. Bacterial inputs to buried sedimentary organic carbon and their impact on deep organic matter maturation processes will be assessed, and the interplay between biological and geological processes in the formation of carbonate mounds can then be defined.
2. Mound "events" frame into a paleoenvironmental plot—prominent erosional surfaces reflect global oceanographic events.
Erosional surfaces are displayed on high-resolution seismic lines. Cores penetrating these unconformities have to be analyzed by means of high-resolution stratigraphy. The well-established nannoplankton and microplankton (planktonic foraminifers) biostratigraphy for the Neogene marine sections of the North Atlantic will support interpretations of unconformities. Oxygen isotope analysis on both planktonic and benthic foraminifers will be run to match isotope data with established orbitally tuned isotope stratigraphy. Absolute ages will be obtained from corals and foraminifers by applying 14C accelerator mass spectrometry (AMS) and U/Th dating for the youngest sequences and 10Be for the older strata. In addition to these tools, absolute ages will be determined on volcanic ashes that occur in the deeper part of the sections. Cores where sufficient biogenic calcite is continuously available will give access to 87Sr/86Sr stratigraphy. All these methods will provide necessary hints for tying the unconformities into a regional and global scheme of oceanographic events.
3. Mounds are high-resolution paleoenvironmental recorders.
To study paleoenvironmental variability a series of well-established proxies will be used. 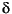 18O analyses will be applied to foraminifers, corals, and other skeletal biota to reconstruct paleotemperatures. As this proxy is also influenced by evaporation/precipitation changes expressed as salinity, we shall run parallel Sr/Ca analyses from aragonitic skeletons to differentiate between temperature and salinity variations. In addition, we may run certain U/Ca analyses, which seem to be an appropriate indicator of paleotemperature as well. For the same reason, we shall use Mg/Ca ratios in calcitic foraminifers. 13C is a useful proxy to reconstruct paleoproductivity; however, it has been demonstrated that it also may indicate methane-derived carbon in carbonates. Therefore, we shall run Ba/Ca analyses to differentiate between productivity signals and methanogenesis. Paleoproductivity can also be determined via 15N in organic matter. Another important question focuses on the carbonate production of the mounds. This can be traced by bulk Sr analyses in combination with total organic carbon (TOC) and CaCO3 determinations in the peri-mound cores. In addition to these geochemical proxies we shall apply classical methods like component analyses and grain size distribution. The sortable silt fraction (10–63 rameter for paleocurrent speed assessment. X-ray radiography will be used to observe delicate structures (ripples and cross stratification), hidden objects (dropstones and coral debris), or bioturbation (Zoophycos burrows) and will allow accurate positioning of subsamples. A promising tool is the application of fluorescence microscopy for organic matter petrography to differentiate between marine-derived material and terrestrial-derived material like leaves and tiny wood fragments. This, in combination with clay mineral analyses, will allow for a distinct differentiation of sediment sources between glacials and interglacials.
18O analyses will be applied to foraminifers, corals, and other skeletal biota to reconstruct paleotemperatures. As this proxy is also influenced by evaporation/precipitation changes expressed as salinity, we shall run parallel Sr/Ca analyses from aragonitic skeletons to differentiate between temperature and salinity variations. In addition, we may run certain U/Ca analyses, which seem to be an appropriate indicator of paleotemperature as well. For the same reason, we shall use Mg/Ca ratios in calcitic foraminifers. 13C is a useful proxy to reconstruct paleoproductivity; however, it has been demonstrated that it also may indicate methane-derived carbon in carbonates. Therefore, we shall run Ba/Ca analyses to differentiate between productivity signals and methanogenesis. Paleoproductivity can also be determined via 15N in organic matter. Another important question focuses on the carbonate production of the mounds. This can be traced by bulk Sr analyses in combination with total organic carbon (TOC) and CaCO3 determinations in the peri-mound cores. In addition to these geochemical proxies we shall apply classical methods like component analyses and grain size distribution. The sortable silt fraction (10–63 rameter for paleocurrent speed assessment. X-ray radiography will be used to observe delicate structures (ripples and cross stratification), hidden objects (dropstones and coral debris), or bioturbation (Zoophycos burrows) and will allow accurate positioning of subsamples. A promising tool is the application of fluorescence microscopy for organic matter petrography to differentiate between marine-derived material and terrestrial-derived material like leaves and tiny wood fragments. This, in combination with clay mineral analyses, will allow for a distinct differentiation of sediment sources between glacials and interglacials.
4. The Porcupine mounds are present-day analogs for Phanerozoic reef mounds and mud mounds.
Mounds are a fundamental and recurrent strategy of life from Precambrian times onward. The true dawn of carbonate mud mounds is in Cambrian times, in the early Paleozoic, when mounds suddenly feature a diversity in microbial and biodetrital fabrics with abundant mound-building calcified microbes, calcified coralline, and green algae and a variety of Paleozoic benthic invertebrates, which may have played an ancillary role in mound construction. In mid- to late Ordovician times, the dramatic rise of large skeletal metazoans such as stromatoporoids, corals (rugosa and tabulata), and bryozoans, as well as higher algae, paved the way for the strong development of reefs and typical stromatactoid mud mounds. Lower Devonian mounds (Gedinnian) in the Montagne Noire exhibit the most spectacular stromatactis fabrics, interpreted as the result of decaying microbial mats. Stratigraphically younger (Emsian) conical carbonate mounds (kess-kess) of the Moroccan Anti-Atlas are related to precipitation from hydrothermal fluids, some of which are inferred to be related to a light carbon source (hydrocarbon). Some of the most impressive of early Carboniferous bank aggregates, reaching up to 1 km in thickness, are those known as the Waulsortian. In full Mesozoic times, a decline in the abundance and diversity of microbial mounds is recorded from the Triassic to the Cretaceous. From the mid-Cretaceous onward, microbial fabrics are only known as components to metazoan framework reefs. Most Cenozoic mud mounds would be of biodetrital origin, although microbial components might have remained significant in deeper water. Drilling Challenger mound will not only allow framing this modern mound within this lineage but also will allow possible identification of the key actors in mound building. To study both skeletal and bacterial fabrics, thin sections of lithified and nonlithified parts of cores will be prepared for comparative studies. In addition, a detailed carbonate petrography shall be applied, including fluorescence microscopy and cathodoluminescence microscopy.
Next Section |
Table of Contents